Неправильный развал схождение последствия: Сход-развал и его влияние на износ шин
Сход-развал и его влияние на износ шин
понедельник, 18 июля 2016 г. 12:27:01 Europe/Moscow
Еще с момента использования карет и конных повозок, колеса на них устанавливались с небольшими отклонениями от продольных и поперечных осей автомобиля. Это делалось для разных целей, к примеру, колеса ставились верней частью наружу, что бы при езде, грязь не попадала на пассажиров, или наоборот, что бы колесо не соскочило с оси при потере нагеля (штырь удерживающий колесо). Кареты уже давно ушли в прошлое, но установка колес с различными наклонами дошла и используется в современных автомобилях. Эти настройки называются углами установки колес. Далее мы рассмотрим основные из них.
Схождение/расхождение колес
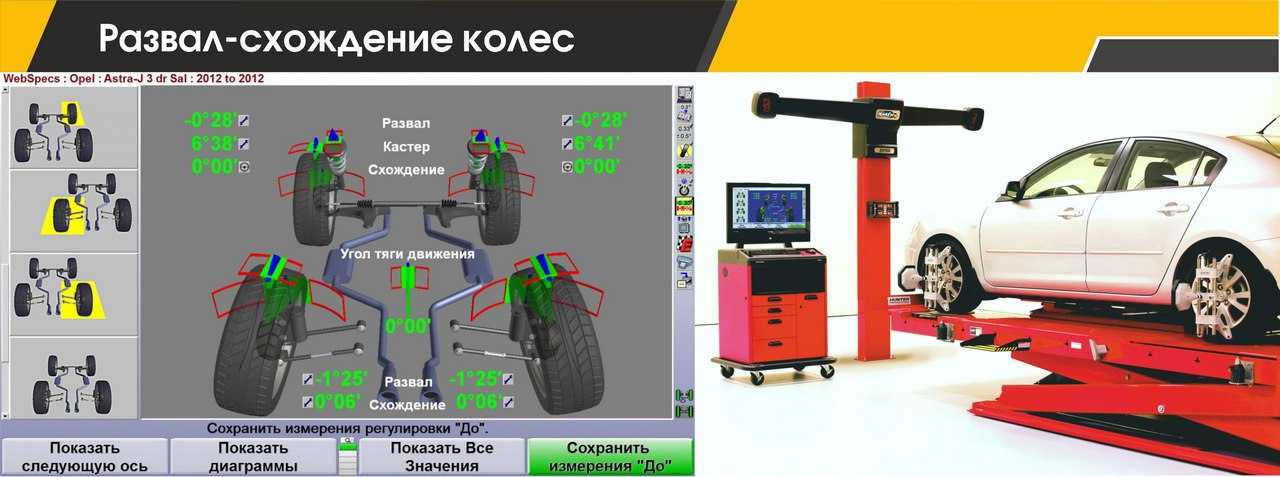
В сервисных центрах мы часто видим предложение отрегулировать сход-развал. В словосочетании «сход-развал» кроются сразу два параметра установки колес. Первым является схождение/расхождение. Схождение представляет собой угол между плоскостью вращения колеса и продольной осью автомобиля при взгляде сверху. Если плоскости колес пересекаются спереди автомобиля (как на картинке справа), то это называется схождением, если позади автомобиля, то расхождением.
Если плоскости колес пересекаются спереди автомобиля (как на картинке справа), то это называется схождением, если позади автомобиля, то расхождением.
Подавляющее большинство серийных автомобилей имеет схождение передних колес. Во-первых, это позволяет компенсировать действие продольных сил на колесо при качении. Во-вторых, это улучшает управляемость и стабильность на дороге. Существуют также обратные примеры: при небольшом расхождении колес увеличивается скорость реакции на поворот руля. Такие настройки часто применяются в автоспорте. Но в повседневной жизни другие преимущества схождения колес намного важнее чрезмерно «острого» руля при расхождении.
Развал колес
Другой важный параметр настройки колес – это развал. Развалом называют угол между вертикалью и плоскостью вращения колеса, если смотреть на автомобиль спереди. Развал может быть положительным (если вершина колеса смотрит наружу) или отрицательным (если колесо смотрит в сторону кузова). С развитием различных типов подвесок на автомобилях встречаются как положительные углы развала, так и отрицательные. Положительный угол призван компенсировать смещение колес при нагрузке на ось (статическая загрузка, или динамическая при разгоне и торможении). Отрицательный развал улучшает сцепление и устойчивость. Машины с отрицательным развалом можно наблюдать в автомобильных гонках, но такие настройки пагубно влияют на срок службы шин.
Положительный угол призван компенсировать смещение колес при нагрузке на ось (статическая загрузка, или динамическая при разгоне и торможении). Отрицательный развал улучшает сцепление и устойчивость. Машины с отрицательным развалом можно наблюдать в автомобильных гонках, но такие настройки пагубно влияют на срок службы шин.
Как правило, значения регулировок углов установки колес указываются диапазонами, в пределах которых можно их выставлять. Даже небольшие изменения этих углов влияют на управляемость и устойчивость автомобиля, ведь колесо должно сохранять оптимальную позицию во всех режимах движения, на любых скоростях и на различных покрытиях. И с помощью настройки кинематики подвески и выставлению этих углов, тот или иной автопроизводитель может придавать различный характер своим моделям.
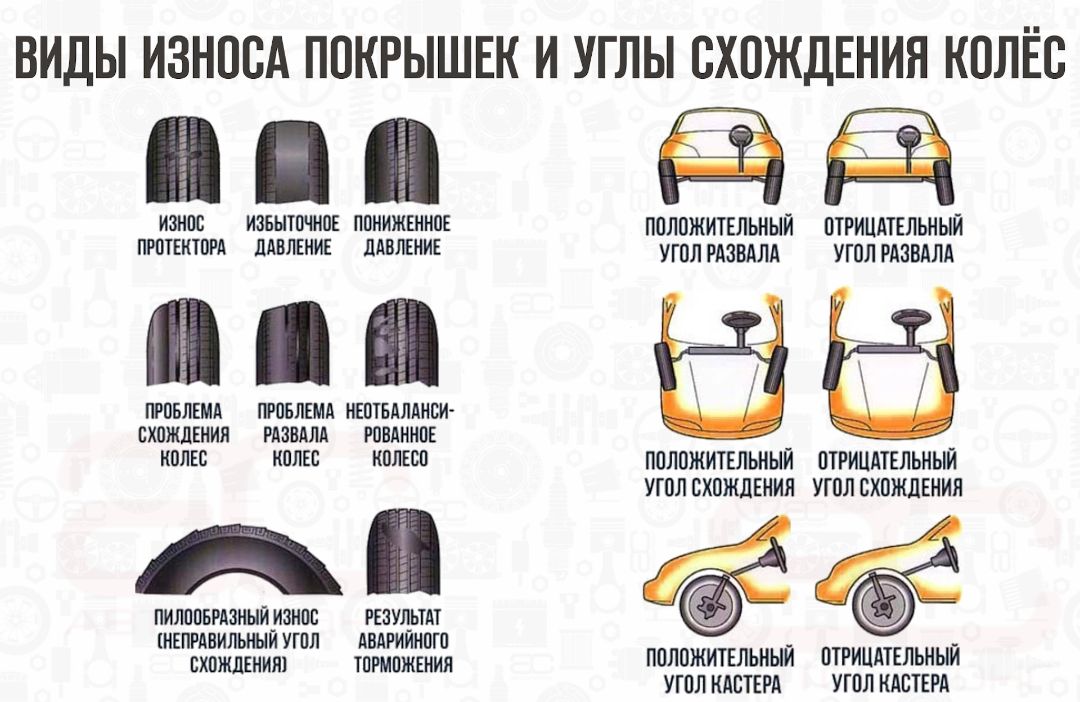
Влияние сход-развала на износ шин
Углы установки колес также влияют на износ шин. При неправильных настройках шина может изнашиваться неравномерно, что выведет её из строя раньше. В некоторых случаях по износу можно определить какие настройки сбились в подвеске.
В некоторых случаях по износу можно определить какие настройки сбились в подвеске.
При неправильном развале будет повышенный износ одной из сторон шины. В случае излишне положительного развала, интенсивнее будет изнашиваться внешняя часть протектора шины. При отрицательном развале, наоборот, сильнее будет изнашиваться внутренняя часть шины. В случае неверных настроек углов схождения колес шина начинает неравномерно стираться и на ней появляются заусенцы, которые можно почувствовать руками. Если вы чувствуете эти заусенцы при движении рукой, от внешней стороны шины к внутренней, то у вас недостаточное схождение колес. А если наоборот, при движении рукой от внутренней части наружу, то схождение колес чрезмерное.
Но это в теории, а в реальности, согласно нашим исследованиям, около 5% летних шин имеют износ увеличивающийся от одного плеча к другому и около 20% имеют изношенное плечо. Всё это происходит из-за неправильных углов установки колес. Поэтому регулярно осматривайте и проверяйте свои шины на предмет повреждений и неравномерного износа и следите за техническим состоянием вашего автомобиля.
Поэтому регулярно осматривайте и проверяйте свои шины на предмет повреждений и неравномерного износа и следите за техническим состоянием вашего автомобиля.
Неправильный угол развал-схождение
Если развал-схождение сделан под неправильным углом, это приведет к «плачевным» результатам в последствии. Это та проблема авто, которая не допускает к себе легкомысленного отношения. Правильный или нет, поставлен угол колес, влияет впоследствии на следующие факты при управлении автомобиля – его управляемость, расход топлива, быстрота изнашивания шин. Человек с немалым опытом вождения, знает, чем оборачивается впоследствии неправильно сделанное развал-схождение. Это аспект необходимо все время держать под контролем, оно не требует больших вложений со стороны владельца.
Итак, в чем же причина нависающих проблем, при некачественной работе авто мастеров? Дело в том, что упругие детали подвески при нагрузке на автомобиль,
начинают прогибаться. Особенно актуально это для тех машин, у которых независимая подвеска. Тяги авто и его рычаги начинают медленно смещаться при проседании
кузова транспортного средства. Это приводит к изменению положения колеса относительно кузова и поверхности асфальта, т.е. усиливается сила трения при движении
авто и влияет на ее маневренность в дальнейшем. Возрастают нагрузки на подвеску и рулевое управление. Урегулировав развал, можно предотвратить нежелательное
воздействие подшипников на ступицу, что увеличит их срок службы.
Тяги авто и его рычаги начинают медленно смещаться при проседании
кузова транспортного средства. Это приводит к изменению положения колеса относительно кузова и поверхности асфальта, т.е. усиливается сила трения при движении
авто и влияет на ее маневренность в дальнейшем. Возрастают нагрузки на подвеску и рулевое управление. Урегулировав развал, можно предотвратить нежелательное
воздействие подшипников на ступицу, что увеличит их срок службы.
- после того, как спидометр показал, что авто прошло 10 т. километров;
- после ремонта ходовой части;
- после «переобувании» колес;
- если замечено неравномерное стирание протекторов;
- нарушен клиренс;
- изменилась маневренность машины, она стала менее управляема;
- руль принимает неправильный оборот при прямом вождении;
- пробита подвеска;
- диски прогнуты и деформированы;
- в целях профилактики – 1 раз в год.
Необходимо уделить пристальное внимание работе по развал — схождению
После этой процедуры, автомобиль становится устойчивым на дороге, легко управляется.
Неправильно сделанный развал — схождения сразу повлечет за собой такие неприятности, как потеря управляемости, увеличиться расход топлива, протектор шин будет претерпевать изменения.
В основном автовладелец замечает непорядок в развале по тому, как начинают изнашиваться шины. Это самая видимая проблема, если угол постановки колес имеет неправильный угол по отношению к поверхности дорожного покрытия. Второе, на что обращает внимание водитель при вождении – это изменение в управления в худшую сторону и ухудшение маневренности авто. Особенно это чувствуется при езде по неровной поверхности — покрышки начинают издавать нехарактерный шум, чувствуется вибрация на руле.
Что следует знать при неисправности генератора
От работы генератора зависит многое в автомобиле – это известно каждому водителю.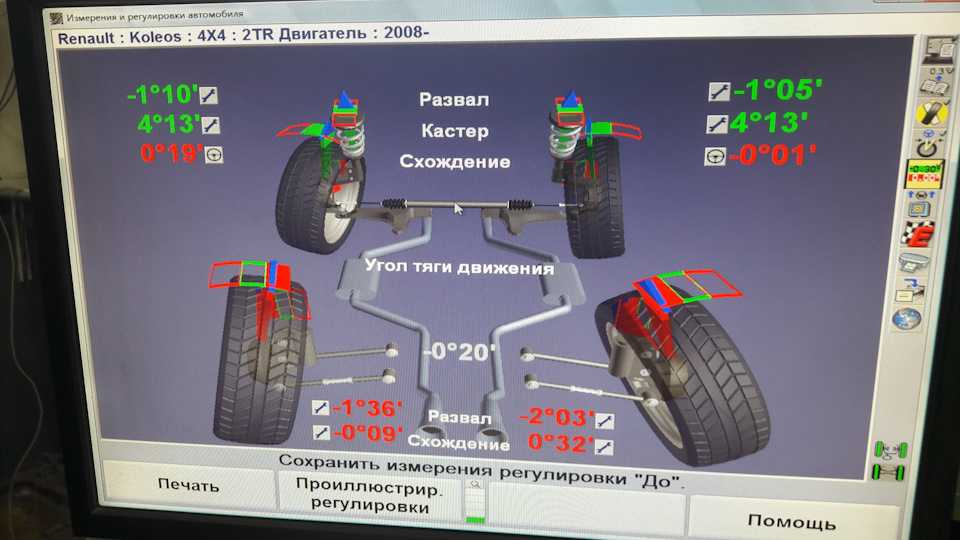 Если взять в расчет электрику машины, то генератор подвергается
наибольшей нагрузке, чем остальные компоненты. Частота оборота в двигателе меньше скорости вращения вала, поэтому неудивительно, что он чаше выходит из
строя, испытывая такие нагрузки. Он бывает переменного тока (его ставят в легковые автомобили) и постоянного тока (применяются в грузовом автотранспорте).
Генератор переменного тока состоит из стартера с неподвижной обмоткой и ротора, создающего магнитное поле. Его преимуществом является скорость вращения
ротора – она высокая. Расстояние со временем меняется между щетками и коллекторными ламелями из-за интенсивной работы. Якорные пазы под воздействием
центробежной силы, выходят из-под обмотки. Щетки с течением времени изнашиваются и требуют замены при достижении высоты в 10 мм. Подшипники подлежат замене,
если роторный вал издает несвойственные посторонние звуки и трудно вращается.
Если взять в расчет электрику машины, то генератор подвергается
наибольшей нагрузке, чем остальные компоненты. Частота оборота в двигателе меньше скорости вращения вала, поэтому неудивительно, что он чаше выходит из
строя, испытывая такие нагрузки. Он бывает переменного тока (его ставят в легковые автомобили) и постоянного тока (применяются в грузовом автотранспорте).
Генератор переменного тока состоит из стартера с неподвижной обмоткой и ротора, создающего магнитное поле. Его преимуществом является скорость вращения
ротора – она высокая. Расстояние со временем меняется между щетками и коллекторными ламелями из-за интенсивной работы. Якорные пазы под воздействием
центробежной силы, выходят из-под обмотки. Щетки с течением времени изнашиваются и требуют замены при достижении высоты в 10 мм. Подшипники подлежат замене,
если роторный вал издает несвойственные посторонние звуки и трудно вращается.
Причины и следствия при неисправности генератора должен знать каждый водитель, даже если он управляет машиной с автоматической коробкой передачи. При ненадлежащей работе генератора – отсутствие тока или неполной подачи зарядки аккумулятору, любому человеку следует знать, какие действия следует
совершить для устранения проблемы, а также причины их возникновения.
При ненадлежащей работе генератора – отсутствие тока или неполной подачи зарядки аккумулятору, любому человеку следует знать, какие действия следует
совершить для устранения проблемы, а также причины их возникновения.
Пробуксовка приводного ремня и повышенный шум, говорит о его недостаточном натяжении. Чтобы устранить это, следует натянуть ремень. Если натяжение не помогло и шум продолжается, следует проверить подшипники.
Если щетки останавливаются, причиной этого может быть грязь, которая налипла на них, мешая работе. В этом случае следует избавиться от нее.
Важно следить за чистотой контактных колец, на них не должно быть масляных капель, иначе они будут пригорать.
Регулятор напряжения не подается ремонту, при неисправности следует его заменить на новый рабочий агрегат.
При неправильной работе генератора, может произойти замыкание и обрыв цепи. Для устранения проблемы, необходимы ремонтные работы в этой части авто.
Если статор начинает «завывать» — это говорит о его неисправности.
 При выходе из строя его нужно заменить на новый.
При выходе из строя его нужно заменить на новый.Со временем гайка шкива может «разболтаться» и привести к неисправности. Проблема решается — простым подтягиванием.
Имеет место в проверке контактов (они не должны быть ржавыми), все провода необходимы быть целыми — без надломов и трещин.
НАШИ УСЛУГИ
Причины и последствия стресса репликации
1. Масаи Х., Мацумото С., Ю З., Йошизава-Сугата Н., Ода М. Репликация ДНК хромосомы эукариот: где, когда и как? Анну. Преподобный Биохим. 2010;79:89–130. [PubMed] [Google Scholar]
2. Woodward A, et al. Избыток Mcm2–7 лицензирует дремлющие ориджины репликации, которые можно использовать в условиях репликативного стресса. Дж. Клеточная биология. 2006; 173: 673–683. [Статья бесплатно PMC] [PubMed] [Google Scholar]
3. Ge X, Jackson D, Blow J. Спящие истоки, лицензированные избытком Mcm2–7, необходимы клеткам человека для выживания в условиях репликативного стресса. Гены Дев. 2007;21:3331–3341. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Макинтош Д., Блоу Дж. Происхождение в состоянии покоя, контрольная точка лицензирования и реакция на репликативный стресс. Харб Колд Спринг. Перспектива. биол. 2012;4 [бесплатная статья PMC] [PubMed] [Google Scholar]
Макинтош Д., Блоу Дж. Происхождение в состоянии покоя, контрольная точка лицензирования и реакция на репликативный стресс. Харб Колд Спринг. Перспектива. биол. 2012;4 [бесплатная статья PMC] [PubMed] [Google Scholar]
5. Pacek M, Walter J. Требование MCM7 и Cdc45 при раскручивании хромосом во время репликации эукариотической ДНК. EMBO J. 2004; 23: 3667–3676. [Статья бесплатно PMC] [PubMed] [Google Scholar]
6. Byun T, Pacek M, Yee M-C, Walter J, Cimprich K. Функциональное разобщение активности хеликазы MCM и ДНК-полимеразы активирует ATR-зависимую контрольную точку. Гены Дев. 2005;19: 1040–1052. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Zou L, Elledge S. Обнаружение повреждения ДНК посредством распознавания ATRIP комплексов RPA-оцДНК. Наука. 2003; 300:1542–1548. [PubMed] [Google Scholar]
8. MacDougall C, Byun T, Van C, Yee M-c, Cimprich K. Структурные детерминанты активации контрольных точек. Гены Дев. 2007; 21: 898–903. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Nam E, Cortez D. Сигнализация ATR: больше, чем встреча на развилке. Биохим. Дж. 2011; 436: 527–536. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Nam E, Cortez D. Сигнализация ATR: больше, чем встреча на развилке. Биохим. Дж. 2011; 436: 527–536. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Марешаль А., Цзоу Л. Определение повреждения ДНК с помощью киназ ATM и ATR. Харб Колд Спринг. Перспектива. биол. 2013;5 [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Bianco J, et al. Анализ профилей репликации ДНК в почкующихся клетках дрожжей и млекопитающих с использованием расчесывания ДНК. Методы. 2012; 57: 149–157. [PubMed] [Google Scholar]
12. Koundrioukoff S, et al. Пошаговая активация сигнального пути ATR при увеличении стресса репликации влияет на целостность хрупкого сайта. Генетика PLoS. 2013;9[Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Ламберт С., Карр А. Препятствия движению репликационной вилки: стабилизация, реактивация и нестабильность генома. Хромосома. 2013; 122:33–45. [PubMed] [Google Scholar]
14. Лабиб К., Де Пикколи Г. Репликация выживших хромосом: множество ролей пути контрольной точки S-фазы.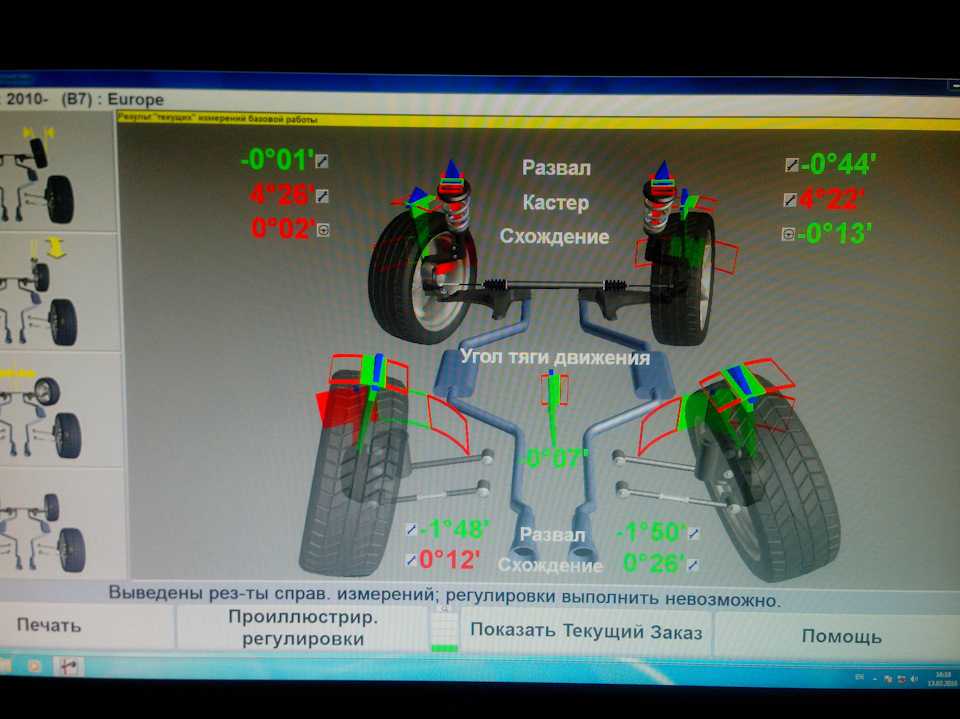 Филос. Транс. Р. Соц. Лонд. Б биол. науч. 2011;366:3554–3561. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Филос. Транс. Р. Соц. Лонд. Б биол. науч. 2011;366:3554–3561. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Petermann E, Helleday T. Пути перезапуска вилки репликации млекопитающих. Нац. Преподобный Мол. Клетка. биол. 2010; 11: 683–687. [PubMed] [Академия Google]
16. Элверс И., Йоханссон Ф., Грот П., Эриксон К., Хелледей Т. Репликационные вилки, остановившиеся в УФ-излучении, перезапускаются путем повторного запуска в фибробластах человека. Нуклеиновые Кислоты Res. 2011;39:7049–7057. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Лопес М., Фояни М., Сого Дж. Несколько механизмов контролируют целостность хромосом после разъединения и перезапуска репликационной вилки при непоправимых УФ-повреждениях. Мол. Клетка. 2006; 21:15–27. [PubMed] [Google Scholar]
18. Mailand N, Gibbs-Seymour I, Bekker-Jensen S. Регуляция взаимодействий PCNA-белок для стабильности генома. Нац. Преподобный Мол. Клетка. биол. 2013;14:269–282. [PubMed] [Google Scholar]
19. Lopes M, et al. Реакция контрольной точки репликации ДНК стабилизирует застопорившиеся вилки репликации. Природа. 2001; 412: 557–561. [PubMed] [Google Scholar]
Lopes M, et al. Реакция контрольной точки репликации ДНК стабилизирует застопорившиеся вилки репликации. Природа. 2001; 412: 557–561. [PubMed] [Google Scholar]
20. Терсеро Дж., Диффли Дж. Регуляция прогрессии вилки репликации ДНК через поврежденную ДНК с помощью контрольной точки Mec1/Rad53. Природа. 2001; 412: 553–557. [PubMed] [Google Scholar]
21. Cobb J, Bjergbaek L, Shimada K, Frei C, Gasser S. Для стабилизации ДНК-полимеразы на остановившихся вилках репликации требуются Mec1 и геликаза RecQ Sgs1. EMBO J. 2003; 22: 4325–4336. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. De Piccoli G, et al. Стабильность реплисом на дефектных вилках репликации ДНК не зависит от киназ контрольной точки S-фазы. Мол. Клетка. 2012; 45: 696–704. [PubMed] [Google Scholar]
23. Ragland R, et al. RNF4 и PLK1 необходимы для коллапса репликационной вилки в ATR-дефицитных клетках. Гены Дев. 2013;27:2259–2273. [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Sirbu BM, et al. Анализ динамики белков на активных, остановленных и свернувшихся вилках репликации. Гены Дев. 2011;25:1320–1327. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Sirbu BM, et al. Анализ динамики белков на активных, остановленных и свернувшихся вилках репликации. Гены Дев. 2011;25:1320–1327. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Hanada K, et al. Структурно-специфическая эндонуклеаза Mus81 способствует перезапуску репликации, генерируя двухцепочечные разрывы ДНК. Нац. Структура Мол. биол. 2007; 14:1096–1104. [PubMed] [Google Scholar]
26. Chanoux R, et al. ATR и h3AX сотрудничают в поддержании стабильности генома в условиях стресса репликации. Дж. Биол. хим. 2009; 284:5994–6003. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Ciccia A, Elledge SJ. Реакция на повреждение ДНК: безопасные игры с ножами. Мол. Клетка. 2010;40:179–204. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Сегурадо М., Диффли Дж. Отдельные роли протеинкиназ контрольной точки повреждения ДНК в стабилизации вилок репликации ДНК. Гены Дев. 2008; 22:1816–1827. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Petermann E, Orta ML, Issaeva N, Schultz N, Helleday T. Репликационные вилки с остановкой гидроксимочевины постепенно инактивируются и требуют двух разных путей, опосредованных RAD51, для перезапуска и Ремонт. Мол. Клетка. 2010;37:492–502. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Petermann E, Orta ML, Issaeva N, Schultz N, Helleday T. Репликационные вилки с остановкой гидроксимочевины постепенно инактивируются и требуют двух разных путей, опосредованных RAD51, для перезапуска и Ремонт. Мол. Клетка. 2010;37:492–502. [Статья бесплатно PMC] [PubMed] [Google Scholar]
30. Матос Дж., Бланко М., Маслен С., Скехель Дж., Уэст С. Регуляторный контроль разрешения промежуточных продуктов рекомбинации ДНК во время мейоза и митоза. Клетка. 2011; 147:158–172. [Статья бесплатно PMC] [PubMed] [Google Scholar]
31. Sørensen C, Syljuåsen R. Защита целостности генома: контрольные киназы ATR, CHK1 и WEE1 сдерживают активность CDK во время нормальной репликации ДНК. Нуклеиновые Кислоты Res. 2012;40:477–486. [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Sogo J, Lopes M, Foiani M. Инверсия вилки и накопление одноцепочечной ДНК в остановленных вилках репликации из-за дефектов контрольных точек. Наука. 2002; 297: 599–602. [PubMed] [Google Scholar]
33. Hu J, et al. Контрольная точка фазы Intra-S нацелена на Dna2, чтобы предотвратить реверсирование остановившихся вилок репликации. Клетка. 2012;149:1221–1232. [PubMed] [Google Scholar]
Hu J, et al. Контрольная точка фазы Intra-S нацелена на Dna2, чтобы предотвратить реверсирование остановившихся вилок репликации. Клетка. 2012;149:1221–1232. [PubMed] [Google Scholar]
34. Couch FB, et al. ATR фосфорилирует SMARCAL1, чтобы предотвратить коллапс вилки репликации. Гены Дев. 2013;27:1610–1623. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Cotta-Ramusino C, et al. Exo1 обрабатывает остановившиеся вилки репликации и противодействует реверсированию вилок в клетках с дефектом контрольной точки. Мол. Клетка. 2005; 17: 153–159. [PubMed] [Google Scholar]
36. Schlacher K, et al. Независимая от репарации двухцепочечного разрыва роль BRCA2 в блокировании деградации остановившейся вилки репликации с помощью MRE11. Клетка. 2011; 145:529–542. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Schlacher K, Wu H, Jasin M. Отдельный путь защиты репликационной вилки соединяет супрессоры опухоли Фанкони с анемией RAD51-BRCA1/2. Раковая клетка. 2012; 22:106–116.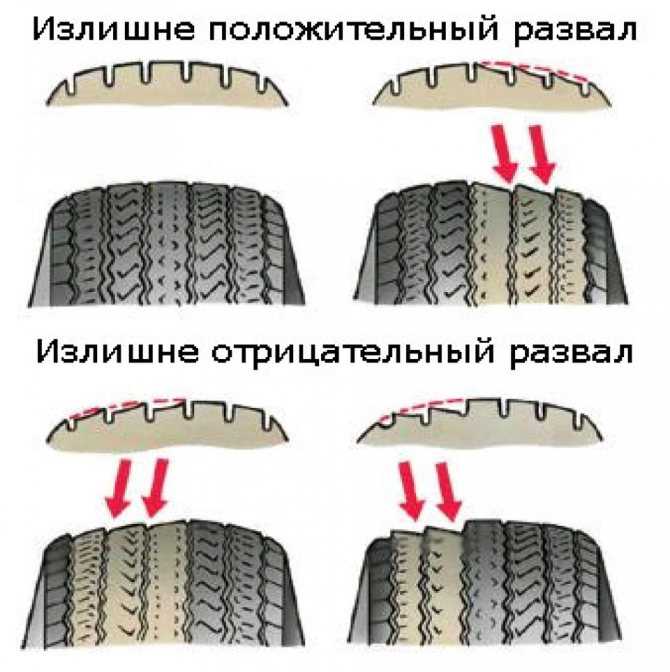 [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Ray Chaudhuri A, et al. Отравление топоизомеразой I приводит к реверсированию вилки репликации, опосредованной PARP. Нац. Структура Мол. биол. 2012;19:417–423. [PubMed] [Google Scholar]
39. Bétous R, et al. SMARCAL1 катализирует регрессию вилки и миграцию соединения Холлидея для поддержания стабильности генома во время репликации ДНК. Гены Дев. 2012;26:151–162. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Shi Y, et al. Деметилирование гистонов, опосредованное гомологом ядерной аминоксидазы LSD1. Клетка. 2004;119: 941–953. [PubMed] [Google Scholar]
41. Брукс П., Терувату Дж. Аддукты ДНК из ацетальдегида: значение для канцерогенеза, связанного с алкоголем. Алкоголь. 2005; 35: 187–193. [PubMed] [Google Scholar]
42. Langevin F, Crossan G, Rosado I, Arends M, Patel K. Fancd2 противодействует токсическому действию альдегидов, вырабатываемых естественным путем у мышей. Природа. 2011; 475:53–58. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
43. Rosado I, Langevin F, Crossan G, Takata M, Patel K. Катаболизм формальдегида необходим в клетках, дефицитных для пути восстановления ДНК при анемии Фанкони. Нац. Структура Мол. биол. 2011;18:1432–1434. [PubMed] [Академия Google]
44. Kim H, D’Andrea A. Регуляция репарации перекрестных связей ДНК с помощью пути анемии Фанкони/BRCA. Гены Дев. 2012; 26:1393–1408. [Статья PMC бесплатно] [PubMed] [Google Scholar]
45. Dalgaard J. Причины и последствия включения рибонуклеотидов в ядерную ДНК. Тенденции Жене. 2012; 28: 592–597. [PubMed] [Google Scholar]
46. Sparks J, et al. Эксцизионная репарация рибонуклеотидов, инициируемая РНКазой h3. Мол. Клетка. 2012;47:980–986. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Reijns M, et al. Ферментативное удаление рибонуклеотидов из ДНК необходимо для целостности и развития генома млекопитающих. Клетка. 2012; 149:1008–1022. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Lazzaro F, et al. РНКаза Н и пострепликационная репарация защищают клетки от рибонуклеотидов, встроенных в ДНК. Мол. Клетка. 2012;45:99–110. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Lazzaro F, et al. РНКаза Н и пострепликационная репарация защищают клетки от рибонуклеотидов, встроенных в ДНК. Мол. Клетка. 2012;45:99–110. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Nick McElhinny S, et al. Нестабильность генома из-за включения рибонуклеотидов в ДНК. Нац. хим. биол. 2010; 6: 774–781. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Kim N, et al. Мутагенный процессинг рибонуклеотидов ДНК дрожжевой топоизомеразой I. Наука. 2011; 332:1561–1564. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Williams J, et al. Опосредованное топоизомеразой 1 удаление рибонуклеотидов из зарождающейся ведущей цепи ДНК. Мол. Клетка. 2013;49:1010–1015. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. McMurray C. Механизмы нестабильности тринуклеотидных повторов в процессе развития человека. Нац. Преподобный Жене. 2010; 11: 786–79.9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Ким Дж., Миркин С. Уравновешивание экспансии повторов ДНК. Курс. мнение Жене. Дев. 2013; 23: 280–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Курс. мнение Жене. Дев. 2013; 23: 280–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Paeschke K, et al. Хеликазы семейства Pif1 подавляют нестабильность генома в мотивах G-квадруплекса. Природа. 2013; 497:458–462. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Бохман М., Паешке К., Закиан В. Вторичные структуры ДНК: стабильность и функция структур G-квадруплекса. Нац. Преподобный Жене. 2012;13:770–780. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Helmrich A, Ballarino M, Nudler E, Tora L. Столкновения транскрипции и репликации, последствия и нестабильность генома. Нац. Структура Мол. биол. 2013;20:412–418. [PubMed] [Google Scholar]
57. Bermejo R, Lai M, Foiani M. Предотвращение репликационного стресса для поддержания стабильности генома: разрешение конфликтов между репликацией и транскрипцией. Мол. Клетка. 2012;45:710–718. [PubMed] [Google Scholar]
58. Barlow J, et al. Выявление хрупких сайтов ранней репликации, которые способствуют нестабильности генома. Клетка. 2013; 152: 620–632. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клетка. 2013; 152: 620–632. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Bermejo R, et al. Контрольная точка репликации защищает стабильность вилки, высвобождая транскрибированные гены из ядерных пор. Клетка. 2011; 146: 233–246. [Статья бесплатно PMC] [PubMed] [Google Scholar]
60. Huertas P, Aguilera A. Котранскрипционно образованные гибриды ДНК: РНК опосредуют нарушение элонгации транскрипции и связанную с транскрипцией рекомбинацию. Мол. Клетка. 2003; 12: 711–721. [PubMed] [Google Scholar]
61. Li X, Manley J. Инактивация фактора сплайсинга белка SR ASF/SF2 приводит к нестабильности генома. Клетка. 2005; 122:365–378. [PubMed] [Академия Google]
62. Paulsen RD, et al. Полногеномный скрининг миРНК выявляет разнообразные клеточные процессы и пути, которые обеспечивают стабильность генома. Мол. Клетка. 2009; 35: 228–239. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Вахба Л., Амон Джереми Д., Кошланд Д., Вуйка-Росс М. РНКаза H и множественные факторы биогенеза РНК сотрудничают, чтобы предотвратить гибриды РНК: ДНК, вызывающие нестабильность генома.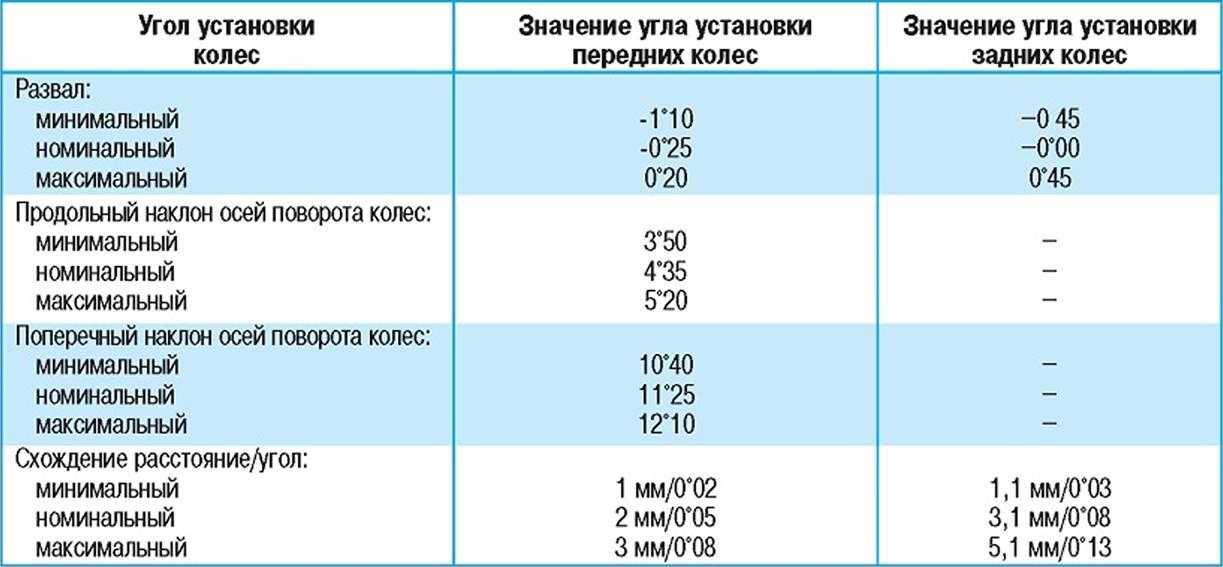 Мол. Клетка. 2011;44:978–988. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Мол. Клетка. 2011;44:978–988. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Stirling P, et al. Опосредованная R-петлей нестабильность генома у мутантов с расщеплением мРНК и полиаденилированием. Гены Дев. 2012; 26: 163–175. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Aguilera A, García-Muse T. Петли R: от побочных продуктов транскрипции до угроз стабильности генома. Мол. Клетка. 2012;46:115–124. [PubMed] [Google Scholar]
66. Tuduri S, et al. Топоизомераза I подавляет нестабильность генома, предотвращая вмешательство между репликацией и транскрипцией. Нац. Клеточная биол. 2009; 11:1315–1324. [Бесплатная статья PMC] [PubMed] [Google Scholar]
67. Bermejo R, et al. Организующие геном факторы Top2 и Hmo1 предотвращают хрупкость хромосом в местах S-фазы транскрипции. Клетка. 2009 г.;138:870–884. [PubMed] [Google Scholar]
68. Alzu A, et al. Сенатаксин связывается с репликационными вилками, чтобы защитить целостность вилок через гены, транскрибированные РНК-полимеразой-II.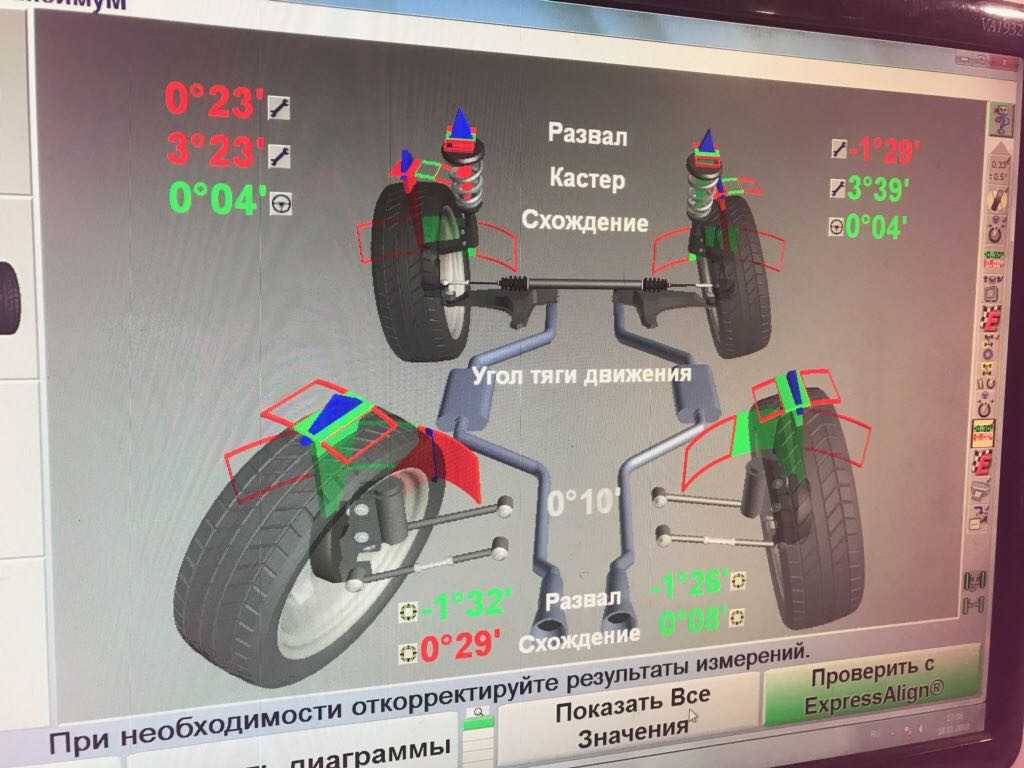 Клетка. 2012; 151:835–846. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Клетка. 2012; 151:835–846. [Бесплатная статья PMC] [PubMed] [Google Scholar]
69. Yüce Ö, West S. Сенатаксин, дефект нейродегенеративного расстройства атаксии с глазодвигательной апраксией 2, лежит на стыке транскрипции и реакции на повреждение ДНК. Мол. Клетка. биол. 2013; 33:406–417. [Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Poli J, et al. Пулы dNTP определяют развитие вилки и использование происхождения в условиях репликационного стресса. EMBO J. 2012; 31: 883–894. [Бесплатная статья PMC] [PubMed] [Google Scholar]
71. Bester A, et al. Дефицит нуклеотидов способствует нестабильности генома на ранних стадиях развития рака. Клетка. 2011; 145:435–446. [Бесплатная статья PMC] [PubMed] [Google Scholar]
72. Англана М., Апиу Ф., Бенсимон А., Дебатисс М. Динамика репликации ДНК в соматических клетках млекопитающих: пул нуклеотидов модулирует выбор источника и расстояние между источниками. Клетка. 2003; 114: 385–39.4. [PubMed] [Google Scholar]
73. Агилера А., Гарсия-Мьюз Т. Причины нестабильности генома. Анну. Преподобный Жене. 2013;47:19–50. [PubMed] [Google Scholar]
74. Saldivar J, et al. Инициация нестабильности генома и предопухолевых процессов за счет потери экспрессии Fhit. Генетика PLoS. 2012;8 [Бесплатная статья PMC] [PubMed] [Google Scholar]
75. Beck H, et al. Подавление циклинзависимой киназы киназой WEE1 защищает геном за счет контроля инициации репликации и потребления нуклеотидов. Мол. Клеточная биол. 2012; 32:4226–4236. [Бесплатная статья PMC] [PubMed] [Google Scholar]
76. Shima N, et al. Жизнеспособный аллель Mcm4 вызывает нестабильность хромосом и аденокарциному молочной железы у мышей. Нац. Жене. 2007; 39: 93–98. [PubMed] [Google Scholar]
77. Debatisse M, Le Tallec B, Letessier A, Dutrillaux B, Brison O. Общие хрупкие сайты: новый взгляд на механизмы нестабильности. Тенденции Жене. 2012; 28:22–32. [PubMed] [Google Scholar]
78. Casper A, Nghiem P, Arlt M, Glover T. ATR регулирует стабильность хрупких сайтов. Клетка. 2002; 111: 779–789. [PubMed] [Академия Google]
79. Le Tallec B, et al. Общее профилирование ломких участков в эпителиальных и эритроидных клетках показывает, что большинство рецидивирующих раковых делеций находятся в ломких участках, содержащих большие гены. Cell Rep. 2013; 4:420–428. [PubMed] [Google Scholar]
80. Ying S, et al. MUS81 способствует экспрессии общего хрупкого сайта. Нац. Клеточная биол. 2013;15:1001–1007. [PubMed] [Google Scholar]
81. Naim V, Wilhelm T, Debatisse M, Rosselli F. ERCC1 и MUS81-EME1 способствуют разделению сестринских хроматид путем процессинга промежуточных продуктов поздней репликации в общих хрупких сайтах во время митоза. Нац. Клеточная биол. 2013;15:1008–1015. [PubMed] [Академия Google]
82. Шринивасан С., Домингес-Сола Д., Ван Л., Хириен О., Готье Дж. Cdc45 является критическим эффектором myc-зависимого стресса репликации ДНК. Cell Rep. 2013; 3:1629–1639. [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Jones R, et al. Повышенная инициация репликации и конфликты с транскрипцией лежат в основе индуцированного Cyclin E стресса репликации. Онкоген. 2012;32:3744–3753. [PubMed] [Google Scholar]
84. Halazonetis T, Gorgoulis V, Bartek J. Модель повреждения ДНК, вызванного онкогенами, для развития рака. Наука. 2008;319: 1352–1355. [PubMed] [Google Scholar]
85. Burrell RA, et al. Стресс репликации связывает структурную и количественную нестабильность раковых хромосом. Природа. 2013; 494:492–496. [Бесплатная статья PMC] [PubMed] [Google Scholar]
86. Neelsen K, Zanini I, Herrador R, Lopes M. Онкогены вызывают генотоксический стресс путем митотического процессинга необычных промежуточных продуктов репликации. Дж. Клеточная биология. 2013; 200:699–708. [Бесплатная статья PMC] [PubMed] [Google Scholar]
87. Jiang Y, et al. Общие ломкие участки характеризуются гипоацетилированием гистонов. Гум. Мол. Жене. 2009 г.;18:4501–4512. [Бесплатная статья PMC] [PubMed] [Google Scholar]
88. Murga M, et al. Мышиная модель ATR-Seckel показывает эмбриональный репликативный стресс и ускоренное старение. Нац. Жене. 2009;41:891–898. [Бесплатная статья PMC] [PubMed] [Google Scholar]
89. Ogi T, et al. Идентификация первого пациента с дефицитом ATRIP и новые мутации в ATR определяют клинический спектр синдрома Секеля ATR-ATRIP. Генетика PLoS. 2012;8:e1002945. [Бесплатная статья PMC] [PubMed] [Google Scholar]
90. О’Дрисколл М., Джегго П. Роль путей реакции на повреждение ДНК в развитии мозга и микроцефалии: понимание человеческих заболеваний. DNA Rep. 2008; 7:1039–1050. [PubMed] [Google Scholar]
91. Duursma AM, Driscoll R, Elias JE, Cimprich KA. Роль комплекса MRN в активации ATR посредством набора TOPBP1. Мол. Клетка. 2013; 50:116–122. [Бесплатная статья PMC] [PubMed] [Google Scholar]
92. Lee J, Dunphy W. Комплекс Mre11-Rad50-Nbs1 (MRN) играет особую роль в активации Chk1 в ответ на остановку вилок репликации. Мол. биол. Клетка. 2013; 24:1343–1353. [Бесплатная статья PMC] [PubMed] [Google Scholar]
93. Shiotani B, et al. Два различных режима активации ATR, организованные Rad17 и Nbs1. Cell Rep. 2013; 3:1651–1662. [Бесплатная статья PMC] [PubMed] [Google Scholar]
94. Stracker T, Petrini J. Комплекс MRE11: начиная с концов. Нац. Преподобный Мол. Клетка. биол. 2011;12:90–103. [Бесплатная статья PMC] [PubMed] [Google Scholar]
95. Crow Y, et al. Мутации в генах, кодирующих субъединицы рибонуклеазы h3, вызывают синдром Айкарди-Гутьера и имитируют врожденную вирусную инфекцию головного мозга. Нац. Жене. 2006;38:910–916. [PubMed] [Google Scholar]
96. Бартек Дж., Мистрик М., Барткова Дж. Пороги передачи сигналов стресса репликации при развитии и лечении рака. Нац. Структура Мол. биол. 2012;19:5–7. [PubMed] [Google Scholar]
97. Schoppy D, et al. Онкогенный стресс повышает чувствительность рака мышей к гипоморфному подавлению ATR. Дж. Клин. Инвестировать. 2012; 122: 241–252. [Бесплатная статья PMC] [PubMed] [Google Scholar]
98. Murga M, et al. Использование индуцированного онкогеном репликативного стресса для селективного уничтожения опухолей, управляемых Myc. Нац. Структура Мол. биол. 2011;18:1331–1335. [Бесплатная статья PMC] [PubMed] [Google Scholar]
99. Gilad O, et al. Сочетание подавления ATR с онкогенным Ras синергически увеличивает нестабильность генома, вызывая синтетическую летальность или онкогенез в зависимости от дозировки. Рак Рез. 2010;70:9693–9702. [Бесплатная статья PMC] [PubMed] [Google Scholar]
100. Ruzankina Y, et al. Задержки регенерации тканей и синтетическая летальность у взрослых мышей после комбинированной делеции Atr и Trp53. Нац. Жене. 2009;41:1144–1149. [Бесплатная статья PMC] [PubMed] [Google Scholar]
101. Лопес-Контрерас А., Гутьеррес-Мартинес П., Спекс Дж., Родриго-Перес С., Фернандес-Капетильо О. Дополнительный аллель Chk1 ограничивает индуцированный онкогеном репликативный стресс и способствует трансформации. Дж. Эксп. Мед. 2012; 209: 455–461. [Бесплатная статья PMC] [PubMed] [Google Scholar]
102. Brown E, Baltimore D. Нарушение ATR приводит к фрагментации хромосом и ранней эмбриональной летальности. Гены Дев. 2000; 14: 397–402. [Бесплатная статья PMC] [PubMed] [Google Scholar]
103. Lam M, Liu Q, Elledge S, Rosen J. Chk1 гаплонедостаточен для множества функций, важных для подавления опухоли. Раковая клетка. 2004; 6: 45–59. [PubMed] [Google Scholar]
104. Джексон С.П., Бартек Дж. Реакция на повреждение ДНК в биологии и болезнях человека. Природа. 2009; 461:1071–1078. [Бесплатная статья PMC] [PubMed] [Google Scholar]
105. Fokas E, et al. Ориентация на ATR в ответ на повреждение ДНК и лечение рака. Лечение рака. Rev. 2013 [PubMed] [Google Scholar]
106. Drusco A, et al. Общие гены-супрессоры опухоли хрупкого участка и соответствующие мышиные модели рака. Дж. Биомед. Биотехнолог. 2011; 2011:984505. [Бесплатная статья PMC] [PubMed] [Google Scholar]
107. Станкевич П., Лупски Дж. Структурная изменчивость генома человека и ее роль в заболевании. Анну. преподобный мед. 2010;61:437–455. [PubMed] [Google Scholar]
108. Arlt M, Wilson T, Glover T. Репликационный стресс и механизмы формирования CNV. Курс. мнение Жене. Дев. 2012;22:204–210. [Бесплатная статья PMC] [PubMed] [Google Scholar]
109. Карр А., Ламберт С. Репликационная стресс-индуцированная нестабильность генома: темная сторона поддержания репликации путем гомологичной рекомбинации. Дж. Мол. биол. 2013 [PubMed] [Академия Google]
110. Hu L, et al. Два пути поддержания репликационной вилки сливают инвертированные повторы для перестройки хромосом. Природа. 2013 [бесплатная статья PMC] [PubMed] [Google Scholar]
111. Follonier C, Oehler J, Herrador R, Lopes M. Повторы GAA, связанные с атаксией Фридрейха, вызывают реверсию репликационной вилки и необычные молекулярные соединения. Нац. Структура Мол. биол. 2013;20:486–494. [PubMed] [Google Scholar]
112. Бернштейн К., Ганглофф С., Ротштейн Р. Геликазы ДНК RecQ в репарации ДНК. Анну. Преподобный Жене. 2010;44:393–417. [Бесплатная статья PMC] [PubMed] [Google Scholar]
113. Chabosseau P, et al. Дисбаланс пиримидинового пула, вызванный дефицитом хеликазы BLM, способствует генетической нестабильности при синдроме Блума. Нац. Комм. 2011;2:368. [PubMed] [Google Scholar]
114. Yuan J, Ghosal G, Chen J. Отжиговая геликаза HARP защищает остановившиеся вилки репликации. Гены Дев. 2009; 23: 2394–2399. [Бесплатная статья PMC] [PubMed] [Google Scholar]
115. Yusufzai T, Kong X, Yokomori K, Kadonaga J. Отжигающая геликаза HARP рекрутируется в сайты репарации ДНК посредством взаимодействия с RPA. Гены Дев. 2009 г.;23:2400–2404. [Бесплатная статья PMC] [PubMed] [Google Scholar]
116. Ciccia A, et al. Белок расстройства SIOD SMARCAL1 представляет собой RPA-взаимодействующий белок, участвующий в перезапуске вилки репликации. Гены Дев. 2009;23:2415–2425. [Бесплатная статья PMC] [PubMed] [Google Scholar]
117. Bansbach C, Bétous R, Lovejoy C, Glick G, Cortez D. Отжиговая геликаза SMARCAL1 поддерживает целостность генома в остановившихся вилках репликации. Гены Дев. 2009;23:2405–2414. [Бесплатная статья PMC] [PubMed] [Google Scholar]
118. Postow L, Woo E, Chait B, Funabiki H. Идентификация SMARCAL1 как компонента реакции на повреждение ДНК. Дж. Биол. хим. 2009; 284:35951–35961. [Бесплатная статья PMC] [PubMed] [Google Scholar]
119. Bétous R, et al. Субстратно-селективная репарация и перезапуск репликационных вилок ДНК-транслоказами. Cell Rep. 2013; 3: 1958–1969. [Бесплатная статья PMC] [PubMed] [Google Scholar]
120. Baradaran-Heravi A, et al. Пенетрантность биаллельных мутаций SMARCAL1 связана с экологическими и генетическими нарушениями экспрессии генов. Гум. Мол. Жене. 2012;21:2572–2587. [Бесплатная статья PMC] [PubMed] [Google Scholar]
121. Лавин М.Ф., Йео А.Дж., Бешерел О.Дж. Сенатаксин защищает геном: последствия для нейродегенерации и других аномалий. Редкие заболевания. 2013;1:e25230. [Бесплатная статья PMC] [PubMed] [Google Scholar]
122. Kawabata T, et al. Спасение от остановленной вилки через спящие точки начала репликации в незатронутой S-фазе способствует правильной сегрегации хромосом и подавлению опухоли. Мол. Клетка. 2011;41:543–553. [Статья бесплатно PMC] [PubMed] [Google Scholar]
123. Хоссейн М., Стиллман Б. Мутации синдрома Мейера-Горлина нарушают ингибирующий домен Orc1 CDK и вызывают редупликацию центросом. Гены Дев. 2012;26:1797–1810. [Бесплатная статья PMC] [PubMed] [Google Scholar]
124. Керцендорфер С., Колнаги Р., Абрамович И., Карпентер Г., О’Дрисколл М. Синдром Мейера-Горлина и синдром Вольфа-Хиршхорна: два нарушения развития, подчеркивающие важность эффективная репликация ДНК для нормального развития и нейрогенеза. DNA Rep. 2013; 12: 637–644. [PubMed] [Google Scholar]
125. Hajdu I, Ciccia A, Lewis S, Elledge S. Кандидат 1 на синдром Вольфа-Хиршхорна участвует в клеточном ответе на повреждение ДНК. проц. Натл. акад. науч. США. 2011;108:13130–13134. [Бесплатная статья PMC] [PubMed] [Google Scholar]
126. Kerzendorfer C, et al. Характеристика функциональных последствий гаплонедостаточности NELF-A (WHSC2) и SLBP позволяет выявить новые клеточные фенотипы при синдроме Вольфа-Хиршхорна. Гум. Мол. Жене. 2012;21:2181–2193. [PubMed] [Google Scholar]
127. Спросите K, et al. Коданин-1, мутировавший при анемическом заболевании CDAI, регулирует функцию Asf1 в снабжении гистонами S-фазы. EMBO J. 2012; 31:2013–2023. [Бесплатная статья PMC] [PubMed] [Google Scholar]
128. Griffith E, et al. Мутации в перицентрине вызывают синдром Секкеля с дефектной ATR-зависимой передачей сигналов о повреждении ДНК. Нац. Жене. 2008; 40: 232–236. [Бесплатная статья PMC] [PubMed] [Google Scholar]
129. Sivasubramaniam S, Sun X, Pan Y-R, Wang S, Lee E. Cep164 представляет собой белок-медиатор, необходимый для поддержания стабильности генома посредством модуляции MDC1, RPA и CHK1. Гены Дев. 2008; 22: 587–600. [Бесплатная статья PMC] [PubMed] [Google Scholar]
130. Chaki M, et al. Захват экзома выявляет мутации ZNF423 и CEP164, связывающие почечную цилиопатию с передачей сигналов в ответ на повреждение ДНК. Клетка. 2012; 150: 533–548. [Бесплатная статья PMC] [PubMed] [Google Scholar]
131. Zhou W, et al. Мутации FAN1 вызывают кариомегалический интерстициальный нефрит, связывая хроническую почечную недостаточность с нарушением репарации повреждений ДНК. Нац. Жене. 2012;44:910–915. [Бесплатная статья PMC] [PubMed] [Google Scholar]
132. Choi H, et al. NEK8 связывает ATR-регулируемую репликационную реакцию на стресс и S-фазную активность CDK с почечными цилиопатиями. Мол. Клетка. 2013;51:423–439. [Бесплатная статья PMC] [PubMed] [Google Scholar]
133. Гонсалес-Суарес И., Гонсало С. Уход за геномом: ламины А-типа сохраняют стабильность генома. Ядро. 2010;1:129–135. [Бесплатная статья PMC] [PubMed] [Google Scholar]
134. Хишида Т., Кубота Ю., Карр А.М., Ивасаки Х. Толерантность, зависящая от пути RAD6–RAD18–RAD5, к хроническому низкодозовому ультрафиолетовому свету. Природа. 2008; 457: 612–615. [PubMed] [Академия Google]
135. Хуанг Д., Пининг Б.Д., Паулович А.Г. Предпочтение безошибочной или подверженной ошибкам репарации после репликации у Saccharomyces cerevisiae, подвергшихся воздействию низких доз метилметансульфоната, зависит от клеточного цикла. Мол. Клетка. биол. 2013;33:1515–1527. [Бесплатная статья PMC] [PubMed] [Google Scholar]
136. Манкури Х., Хаттнер Д., Хиксон И. Как незавершенные дела S-фазы влияют на митоз и далее. EMBO J. 2013; 32: 2661–2671. [Бесплатная статья PMC] [PubMed] [Google Scholar]
137. Негрини С., Горгулис В., Халазонетис Т. Геномная нестабильность — развивающийся признак рака. Нац. Преподобный Мол. Клетка. биол. 2010;11:220–228. [PubMed] [Академия Google]
138. Хильдебрандт Ф., Бензинг Т., Кацанис Н. Цилиопатии. Н. англ. Дж. Мед. 2011; 364:1533–1543. [Бесплатная статья PMC] [PubMed] [Google Scholar]
139. Broers J, Hutchison C, Ramaekers F. Laminopathies. Дж. Патол. 2004; 204: 478–488. [PubMed] [Google Scholar]
Помогите! У меня есть предупреждения о конвергенции — biologyforfun
R и Stat
После всей тяжелой работы по сбору данных, обдумыванию соответствующих моделей, форматированию данных, вы, наконец, запускаете свою модель, вот и все, вы получите долгожданные результаты, и БУМ вы получите такое сообщение:
## Предупреждение в checkConv(attr(opt, "derivs"), opt$par, ctrl = ## control$checkConv, : Модель не сошлась с max|grad| = 1,52673 ## (tol = 0,001, компонент 17)
Что это значит? Это плохо? И что вы можете с этим поделать?
Что такое конвергенция?
Лишь немногие модели имеют точное решение, которое может быть автоматически получено из данных, одним из таких случаев является простая линейная модель (соответствующая lm в R). Для этих моделей есть уравнения, дающие все необходимые вам ответы, такие как оценки параметров, p-значения и т. д., основанные только на имеющихся данных (см. эту вики-страницу). Для всех других моделей, таких как GLM или GLMM, необходимо запустить алгоритм, чтобы найти интересующие оценки. По сути, это можно сравнить с просьбой к слепой собаке (алгоритму) найти максимально возможную точку (максимальную вероятность) в определенном ландшафте (пространство параметров) и за ограниченное время (количество итераций). В некоторых случаях, когда есть четкий склон и только одна горка, собака может найти цель всего за несколько итераций. В других случаях, когда вероятный ландшафт состоит из нескольких холмов с большими плоскими участками между этими холмами, бедная слепая собака будет бегать вокруг, но по истечении времени она сообщит вам, что не смогла найти самую высокую возможную точку и выдаст вам предупреждение о сходимости.
Это плохо?
Короче да, очень. Вы не можете доверять оценкам параметров из моделей, которые не сошлись, и тем более другим производным величинам, таким как стандартные ошибки или p-значения. Таким образом, нет необходимости выполнять проверки остатков или отображать результаты, вы должны попытать счастья еще раз, этому режиму нельзя доверять. При этом разработчик пакета должен установить пороговые значения (иногда также называемые допуском), отделяющие модель, которая сходится, от модели, которая не сходится. Обычно разработчик склонен быть более консервативным, чтобы быть уверенным в том, что выводам, сделанным на основе моделей, можно доверять. Но иногда имеющаяся модель очень близка к порогу, например, скажем, что конвергенция отслеживания метрики составляет 2e-6, а порог равен 1e-6, в этом случае не должно быть слишком сложно достичь сходимости небольшими изменениями.
Что я могу сделать?
Ниже приведен неполный список того, что можно сделать, чтобы попытаться достичь сходимости для проблемных моделей, я упорядочил этот список на основе шагов, которые я обычно выполняю при возникновении проблем с сходимостью, вы также можете проверить эту страницу с дополнительными информация для проблем сходимости с lme4:
- стандартизировать все предикторы : если независимые переменные в моделях имеют очень разные масштабы, например, одна переменная идет от -100 до 10000, а другая от 0,001 до 0,01, это делает параметр пространство очень сложное для навигации по алгоритму.
 При стандартизации предикторов все находится в одном масштабе, см. эту статью [ссылка] для получения дополнительных причин для стандартизации ваших предикторов.
При стандартизации предикторов все находится в одном масштабе, см. эту статью [ссылка] для получения дополнительных причин для стандартизации ваших предикторов.
- Попробуйте разные распределения : есть ли смысл попробовать распределение Пуассона? Ваши данные избыточны или недостаточно рассредоточены? Обычно у нас есть довольно хорошее представление о том, какому распределению должны следовать данные, но предупреждения о сходимости можно смягчить либо путем использования более простых распределений, таких как использование распределения Гаусса для аппроксимации распределенных по Пуассону данных, когда среднее значение велико (скажем, больше 10). Кроме того, иногда подобранное распределение ожидает определенных отношений среднего и дисперсии, которые могут быть просто неправильными в имеющихся данных, поэтому попытка более общих распределений, таких как представленный в пакете glmmTMB, может решить проблему сходимости.
- перезапустите модель с оценки параметров, достигнутой до того, как алгоритм сдался : по умолчанию алгоритм начинает со случайных значений параметров, которые могут быть довольно далеки от точки максимальной вероятности. Таким образом, идея состоит в том, чтобы использовать значения параметров из предыдущей неудачной подгонки в качестве отправных точек, в lme4 это делается с помощью следующего кода:
сс <- getME(model_1,c("тета","fixef"))
model_2 <- обновить(model_1,start=ss) - глубоко подумайте о сложности модели : в наши дни так легко подобрать очень сложные модели, не осознавая ослепительной сложности, скрытой под капотом. Сделайте шаг назад и подумайте: действительно ли вам нужны эти вложенные случайные эффекты? Можете ли вы отказаться от этого пятистороннего взаимодействия? Поймите, что модель, которую вы можете подобрать, зависит от количества и качества ваших данных. Вот почему обычно лучше подумать о модели, которую вы будете использовать при разработке сбора данных. С помощью протокола, которому вы будете следовать, сможете ли вы подогнать нужные модели и извлечь сигнал, который вам нужен?
- попробуйте другой алгоритм : обычно для определенного типа модели доступно более одного алгоритма, чтобы получить ответ, например, lme4 предлагает по умолчанию 2 алгоритма Nelder-Mead и bobyqa
- запустить алгоритм с большим количеством итераций : если решение не было найдено в первой 1000 итераций алгоритма, возможно, 10000 будет работать лучше. Но будьте осторожны со временем работы модели, вы же не хотите ждать неделями.
- Соберите больше данных : сложность модели должна расти с размером выборки и/или качеством данных, если в ваших данных есть только неопределенный сигнал, алгоритму будет довольно сложно ориентироваться по поверхности правдоподобия. Более высокое отношение сигнал/шум поможет
- Go Bayesian : модели, оснащенные классической статистикой, используют только информацию из данных, поэтому это означает, что алгоритм получает только указания относительно того, куда он должен пойти, чтобы найти точку наибольшего правдоподобия из данных. Тем не менее, в большинстве случаев мы могли бы предоставить некоторые дополнительные указания алгоритму в виде априорной информации о вероятных значениях параметра модели. Например, в логистической регрессии со стандартизированными предикторами маловероятно, что параметр наклона будет меньше -5 или больше 5 (запустите «curve(invlogit(5*x),-2,2)» в R после загрузки пакет рук). В байесовском анализе данных используются слабоинформативные априорные значения, чтобы предотвратить блуждание алгоритма в пространствах параметров, где значения не имеют большого смысла, но все же оставляют данные, управляющие выборкой. Итак, суть такова: если вы хотите сопоставить сложные модели с небольшим количеством данных, вам может потребоваться предоставить некоторую дополнительную информацию для вашего алгоритма, и байесовская структура именно это и делает (с некоторыми дополнительными тонкостями, которые вы обнаружите по пути). ).
Некоторые ссылки:
Две ссылки, которые очень помогли мне понять концепцию конвергенции и то, как работает алгоритм:
- Книгу экологических моделей Бена Болкера необходимо прочитать, глава 7 по оптимизации особенно актуальна для понимания конвергенции.
- Ричард МакЭлрит Книга «Статистическое переосмысление» великолепна, она намного шире, чем тема этого поста, но чтение первых нескольких глав изменит ваше понимание моделей.
Comments |0|
Category: Разное